
Overexpression of Douglas-Fir LEAFY COTYLEDON1 (PmLEC1) in Arabidopsis Induces Embryonic Programs and Embryo-like Structures in the lec1-1 Mutant but Not in Wild Type Plants


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
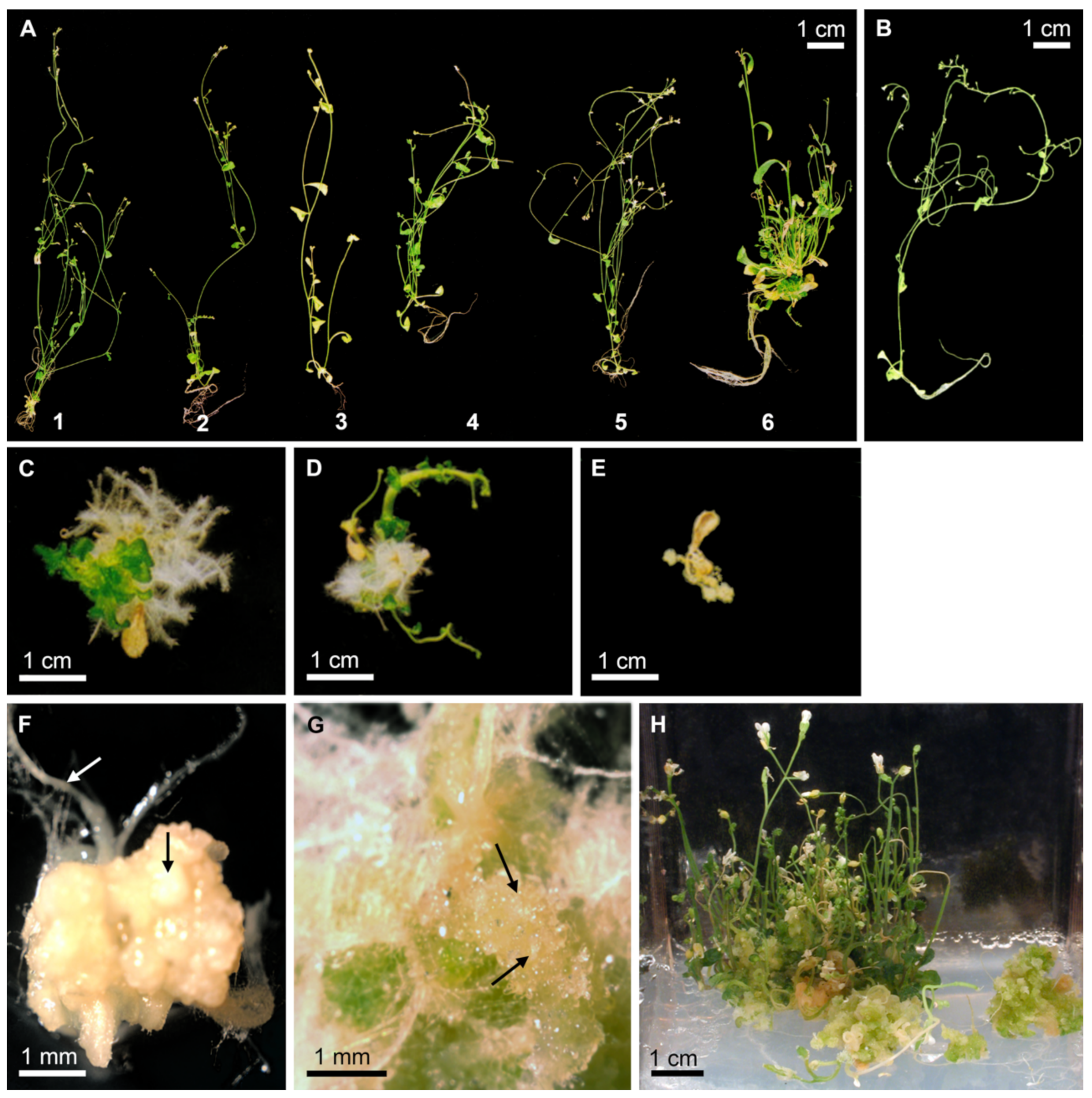
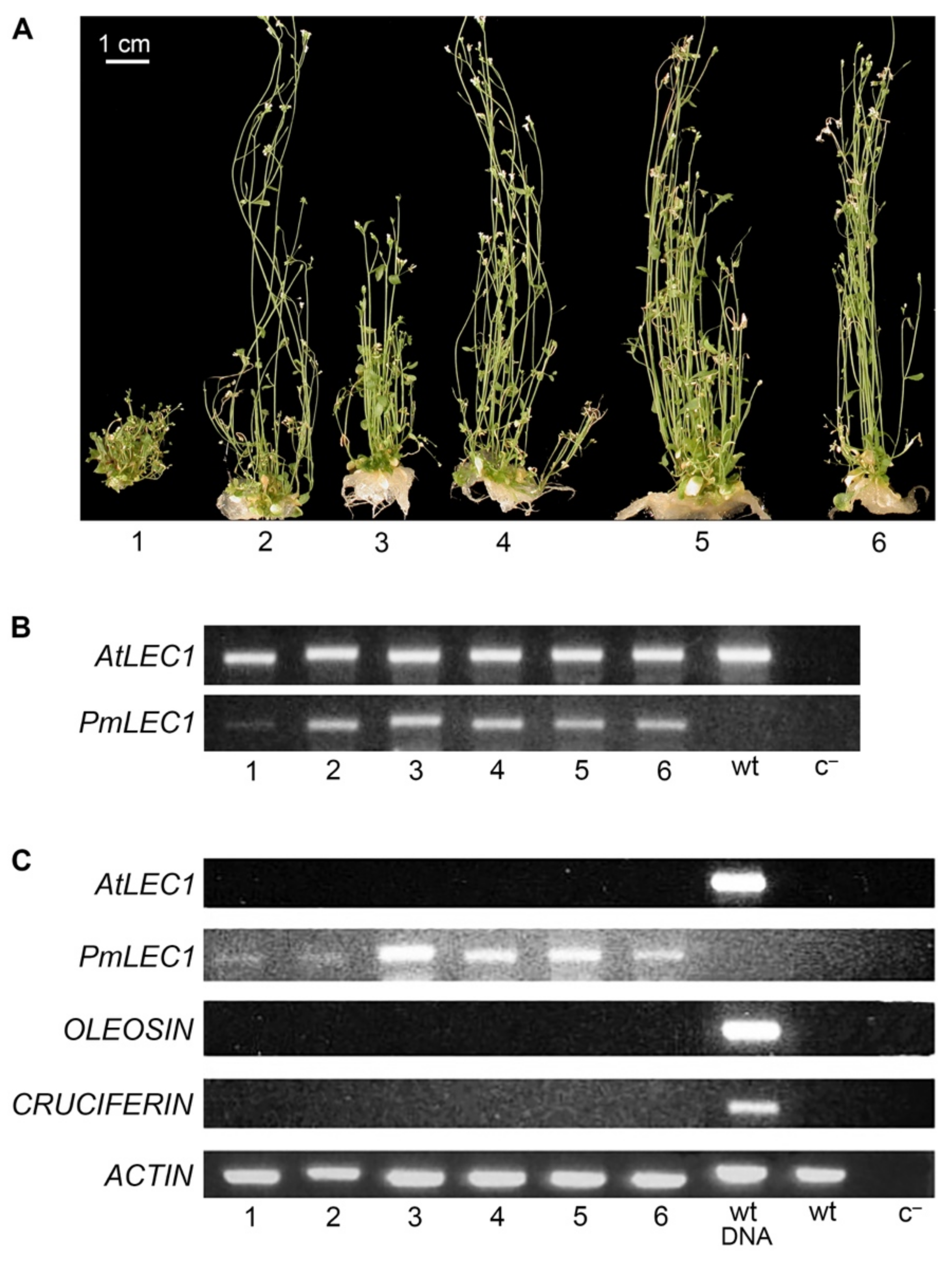
2.1. PmLEC1 Rescues the lec1-1 Null Mutant, Inhibits Vegetative Development and Leads to the Formation of Embryo-like Structures in Abnormal T1 Seedlings
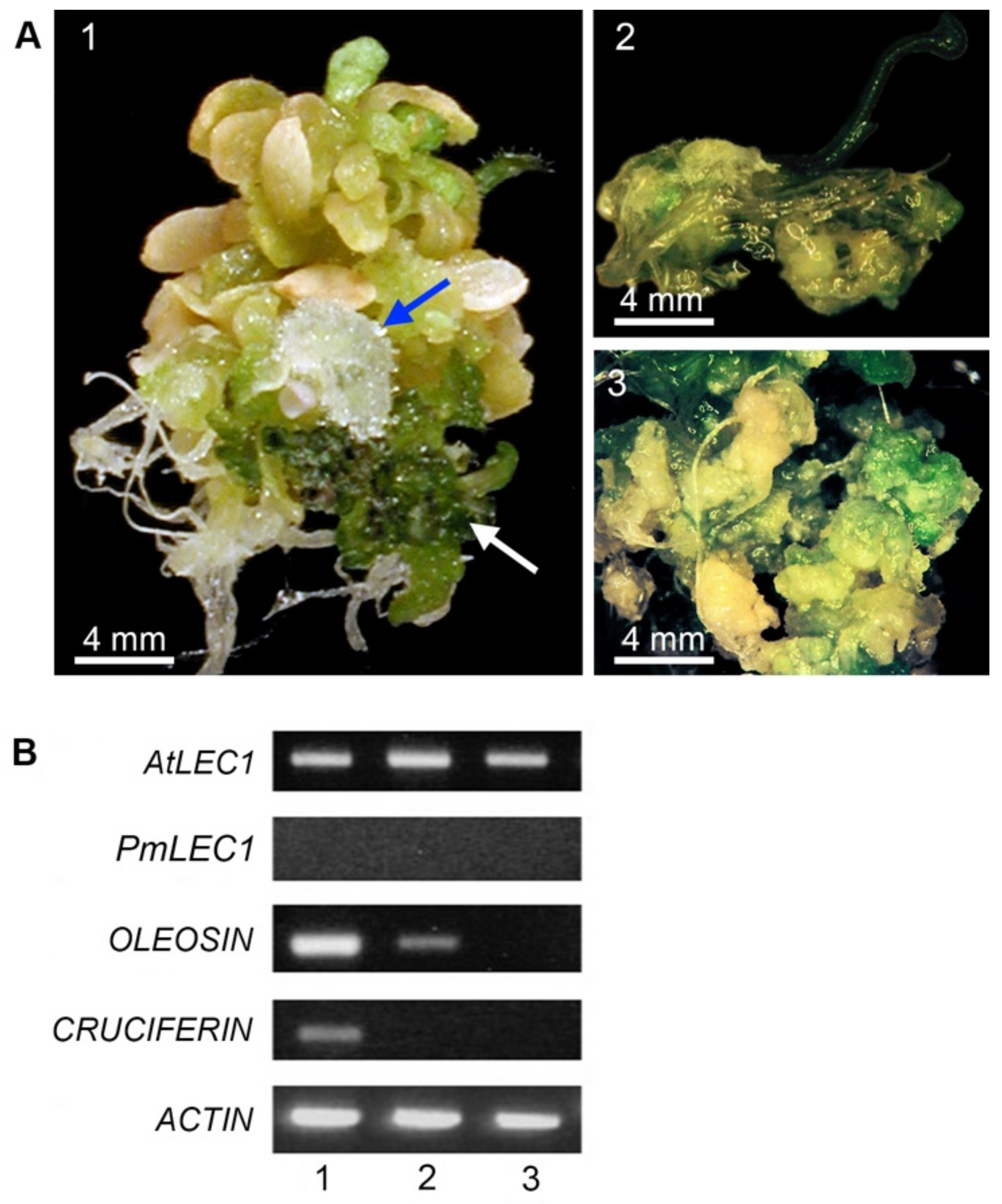
2.2. Ectopic Expression of PmLEC1 in the lec1-1PmLEC1 T2 Generation Induces Embryonic Programs and Spontaneous Formation of Embryo-like Structures
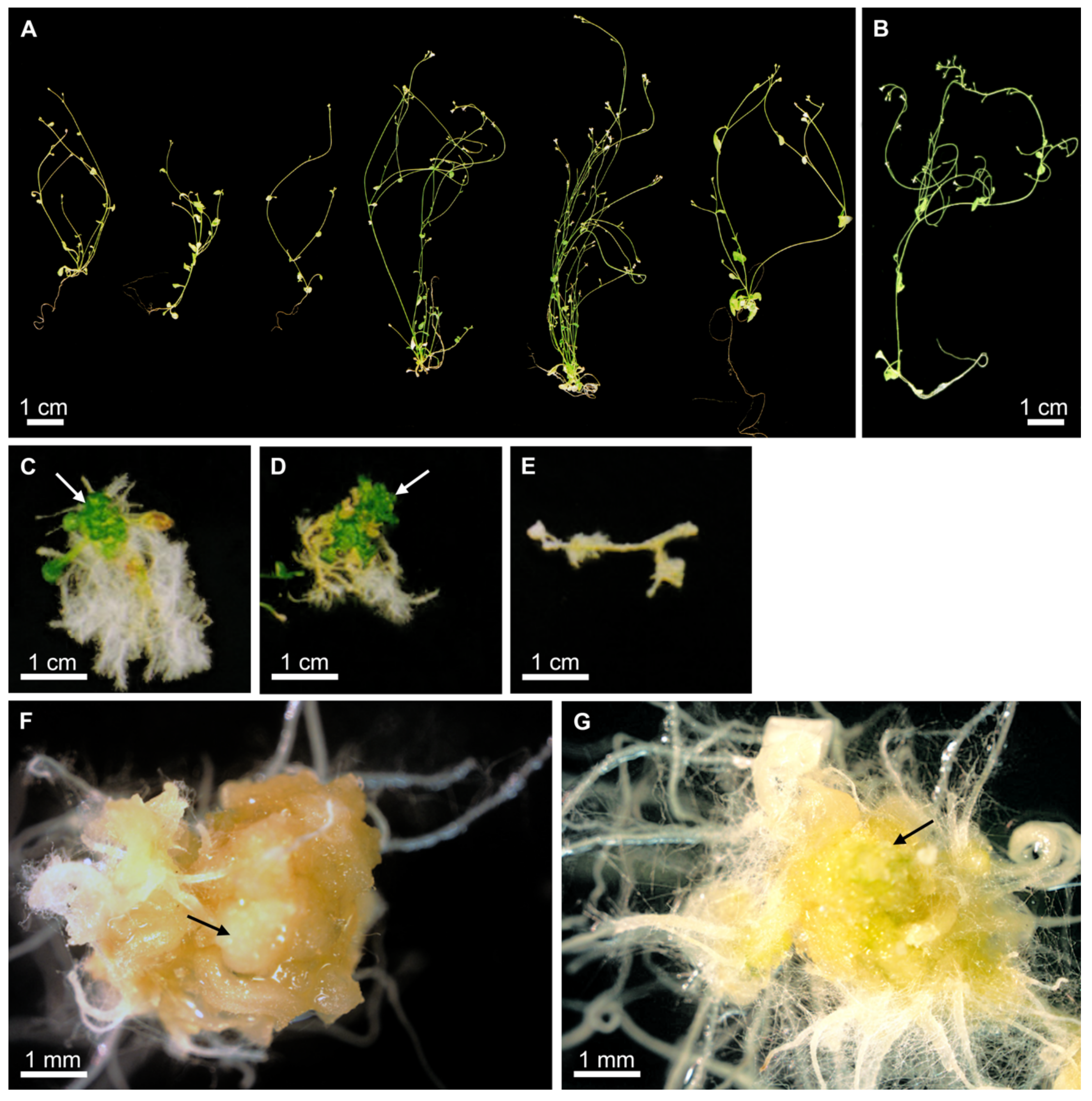
2.3. Overexpression of PmLEC1 in Wild Type Plants Reduces Apical Dominance but Does Not Activate Embryonic Programs
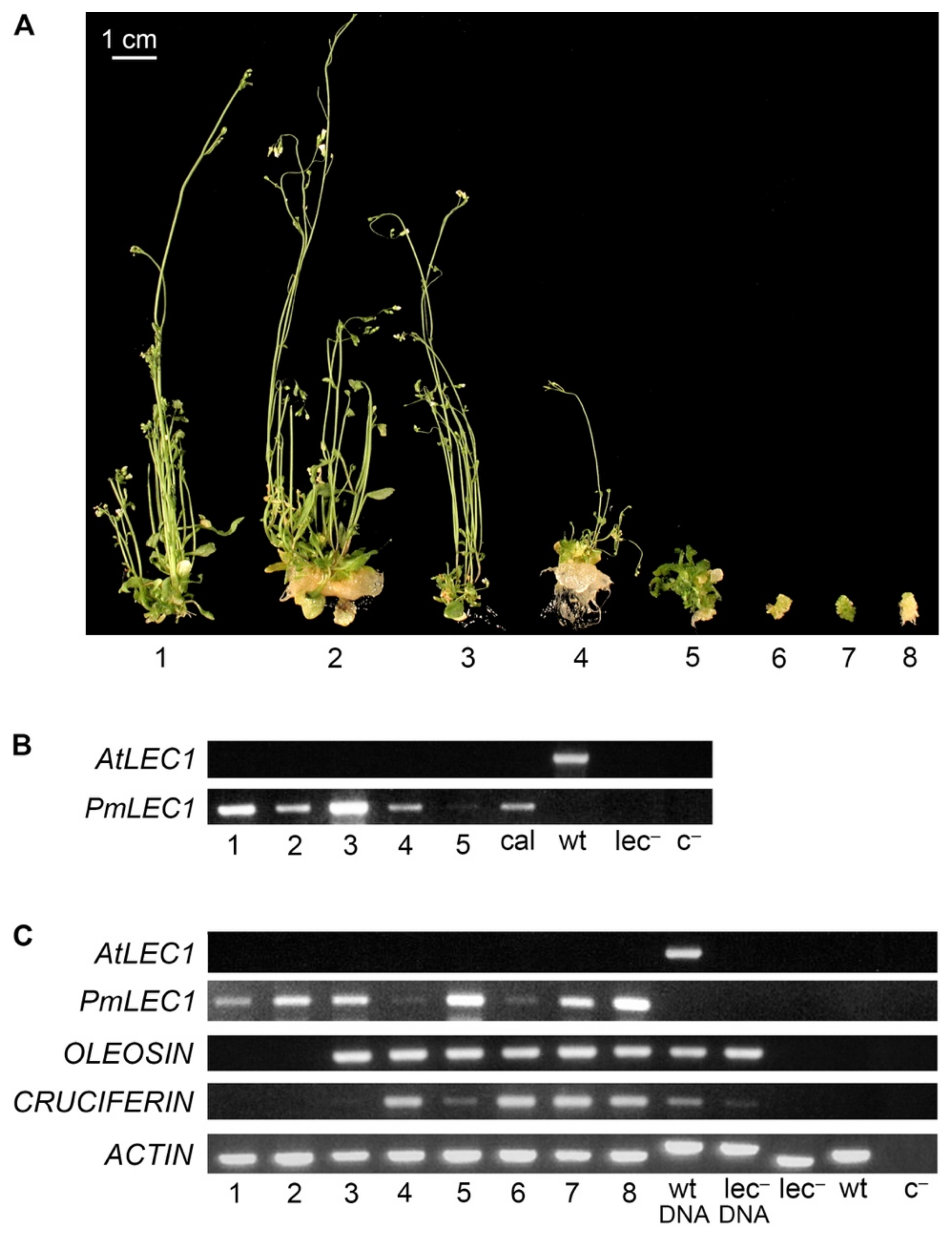
2.4. PmLEC1 Protein Accumulation in Transgenic Plants Correlates with Embryo-like Character and Suppression of Vegetative Development
3. Discussion
3.1. Douglas-Fir PmLEC1 Is Functionally Homologous to Arabidopsis LEC1 and Induces Embryonic Programs in the Arabidopsis lec1-1 Null Mutant
3.2. Transgene Silencing May Account for the Episodes of Normal Development Observed in lec1-1PmLEC1 T1 and T2 Transgenic Plants
3.3. PmLEC1 May Function Prior to Embryogenesis
3.4. LEC1 by Itself Cannot Induce De Novo Embryogenesis
3.5. The AtLEC1 Promoter Region May Exert Control over Embryogenesis
4. Materials and Methods
4.1. Plant Material
4.2. Construction of Expression Cassettes
4.3. Agrobacterium-Mediated Plant Transformation
4.4. DNA Isolation and PCR Analysis
4.5. RNA Isolation and RT-PCR Analysis
4.6. Western Blot Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cairney, J.; Xu, N.; MacKay, J.; Pullman, J. Transcript profiling: A tool to assess the development of conifer embryos. In Vitro Cell. Dev. Biol. Plant 2000, 36, 155–162. [Google Scholar] [CrossRef]
- Lotan, T.; Ohto, M.; Yee, K.M.; West, M.A.; Lo, R.; Kwong, R.W.; Yamagishi, K.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. Arabidopsis LEAFY COTYLEDON1 is sufficient to induce embryo development in vegetative cells. Cell 1998, 93, 1195–1205. [Google Scholar] [CrossRef] [Green Version]
- West, M.A.L.; Yee, K.M.; Danao, J.; Zimmerman, J.L.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON1 Is an Essential Regulator of Late Embryogenesis and Cotyledon Identity in Arabidopsis. Plant Cell 1994, 6, 1731–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinke, D.W.; Franzmann, L.H.; Nickle, T.C.; Yeung, E.C. Leafy Cotyledon Mutants of Arabidopsis. Plant Cell 1994, 6, 1049–1064. [Google Scholar] [CrossRef] [PubMed]
- Gaj, M.D.; Zhang, S.; Harada, J.J.; Lemaux, P.G. Leafy cotyledon genes are essential for induction of somatic embryogenesis of Arabidopsis. Planta 2005, 222, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Vetrici, M.A.; Yevtushenko, D.P.; Misra, S. Douglas-fir LEAFY COTYLEDON1 (PmLEC1) is an active transcription factor during zygotic and somatic embryogenesis. Plant Direct 2021, 5, e00333. [Google Scholar]
- Kwong, R.W.; Bui, A.Q.; Lee, H.; Kwong, L.W.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON1-LIKE defines a class of regulators essential for embryo development. Plant Cell 2003, 15, 5–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datla, R.S.S.; Bekkaoui, F.; Hamerlindl, J.K.; Pilate, G.; Dunstan, D.I.; Crosby, W.L. Improved high-level constitutive foreign gene expression in plants using an AMV RNA4 untranslated leader sequence. Plant Sci. 1993, 94, 139–149. [Google Scholar] [CrossRef]
- Mordhorst, A.P.; Voerman, K.J.; Hartog, M.V.; Meijer, E.A.; van Went, J.; Koornneef, M.; de Vries, S.C. Somatic embryogenesis in Arabidopsis thaliana is facilitated by mutations in genes repressing meristematic cell divisions. Genetics 1998, 149, 549–563. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Koop, H.-U. Somatic embryogenesis in cultured immature zygotic embryos and leaf protoplasts of Arabidopsis thaliana ecotypes. Planta 1997, 202, 387–396. [Google Scholar] [CrossRef]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.; Kohli, A.; Twyman, R.M.; Christou, P. Alternative silencing effects involve distinct types of non-spreading cytosine methylation at a three-gene, single-copy transgenic locus in rice. Mol. Gen. Genet. 2000, 263, 106–118. [Google Scholar] [CrossRef] [PubMed]
- De Wilde, C.; Van Houdt, H.; De Buck, S.; Angenon, G.; De Jaeger, G.; Depickler, A. Plants as bioreactors for protein production: Avoiding the problem of transgene silencing. Plant Mol. Biol. 2000, 43, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Minocha, S.C. Optimization of the expression of a transgene in plants. In Molecular Biology of Woody Plants; Mohan Jain, S., Minocha, S.C., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; Volume 1, pp. 1–30. [Google Scholar]
- Matzke, M.A.; Matzke, A.J.M. How and why do plants inactivate homologous (trans) genes? Plant Physiol. 1995, 107, 679–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yevtushenko, D.P.; Misra, S. Comparison of pathogen-induced expression and efficacy of two amphibian antimicrobial peptides, MsrA2 and temporin A, for engineering wide-spectrum disease resistance in tobacco. Plant Biotechnol. J. 2007, 5, 720–734. [Google Scholar] [CrossRef] [Green Version]
- Holmquist, G.P.; Ashely, T. Chromosome organization and chromatin modification: Influence on genome function and evolution. Cytogenet. Genome Res. 2006, 114, 96–125. [Google Scholar] [CrossRef]
- Grishok, A.; Sinskey, J.L.; Sharp, P.A. Transcriptional silencing of a transgene by RNAi in the soma of C. elegans. Genes Dev. 2005, 19, 683–696. [Google Scholar] [CrossRef] [Green Version]
- Casson, S.A.; Lindsey, K. The turnip mutant of Arabidopsis reveals that LEAFY COTYLEDON1 expression mediates the effects of auxin and sugars to promote embryonic cell identity. Plant Physiol. 2006, 142, 26–41. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Z.; Santes, C.M.; Engel, M.L.; Gasser, C.S.; Harada, J.J. DNA sequences that activate isocitrate lyase gene expression during late embryogenesis and during postgerminative growth. Plant Physiol. 1996, 110, 1069–1079. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 7, 248–254. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vetrici, M.A.; Yevtushenko, D.P.; Misra, S. Overexpression of Douglas-Fir LEAFY COTYLEDON1 (PmLEC1) in Arabidopsis Induces Embryonic Programs and Embryo-like Structures in the lec1-1 Mutant but Not in Wild Type Plants. Plants 2021, 10, 1526. https://doi.org/10.3390/plants10081526
Vetrici MA, Yevtushenko DP, Misra S. Overexpression of Douglas-Fir LEAFY COTYLEDON1 (PmLEC1) in Arabidopsis Induces Embryonic Programs and Embryo-like Structures in the lec1-1 Mutant but Not in Wild Type Plants. Plants. 2021; 10(8):1526. https://doi.org/10.3390/plants10081526
Chicago/Turabian StyleVetrici, Mariana A., Dmytro P. Yevtushenko, and Santosh Misra. 2021. "Overexpression of Douglas-Fir LEAFY COTYLEDON1 (PmLEC1) in Arabidopsis Induces Embryonic Programs and Embryo-like Structures in the lec1-1 Mutant but Not in Wild Type Plants" Plants 10, no. 8: 1526. https://doi.org/10.3390/plants10081526